(UroToday.com) The Bladder Cancer Advocacy Network (BCAN) Bladder Cancer Think Tank 2025, held in Washington D.C., United States, was host to the session: BCAN 2023 Bladder Cancer Research Innovation Awardees. Dr Christopher Garris discussed how to overcome immune suppressive myeloid cell barriers in bladder cancer.
Dr. Garris began his presentation by revisiting a foundational concept in tumor immunology: the Cancer Immunity Cycle. He emphasized that dendritic cells (DCs) play a central role in orchestrating antitumor immune responses by guiding the activation and function of T cells. The cycle begins with the release of tumor antigens following cancer cell death, which are then captured and presented by antigen-presenting cells, particularly dendritic cells.1 This antigen presentation in lymph nodes leads to the priming and activation of T cells, which subsequently traffic to the tumor microenvironment, recognize cancer cells, and ultimately execute cytotoxic killing as illustrated below.

Furthermore, Dr. Garris reviewed the fundamental three-signal model of dendritic cell (DC)–mediated T cell activation, a cornerstone of effective antitumor immunity. He highlighted how this tightly regulated sequence ensures both specificity and potency in the adaptive immune response. Signal 1 involves antigen presentation: the T cell receptor (TCR) on CD4⁺ or CD8⁺ T cells recognizes a specific antigen (Ag) presented by major histocompatibility complex (MHC) molecules on the surface of antigen-presenting cells (APCs), typically dendritic cells. Signal 2 is the co-stimulatory interaction, often mediated by CD28 on the T cell engaging with CD80/CD86 on the APC. Finally, Signal 3 encompasses the release of cytokines by the APC, which shape T cell differentiation and functional polarization as illustrated below.1

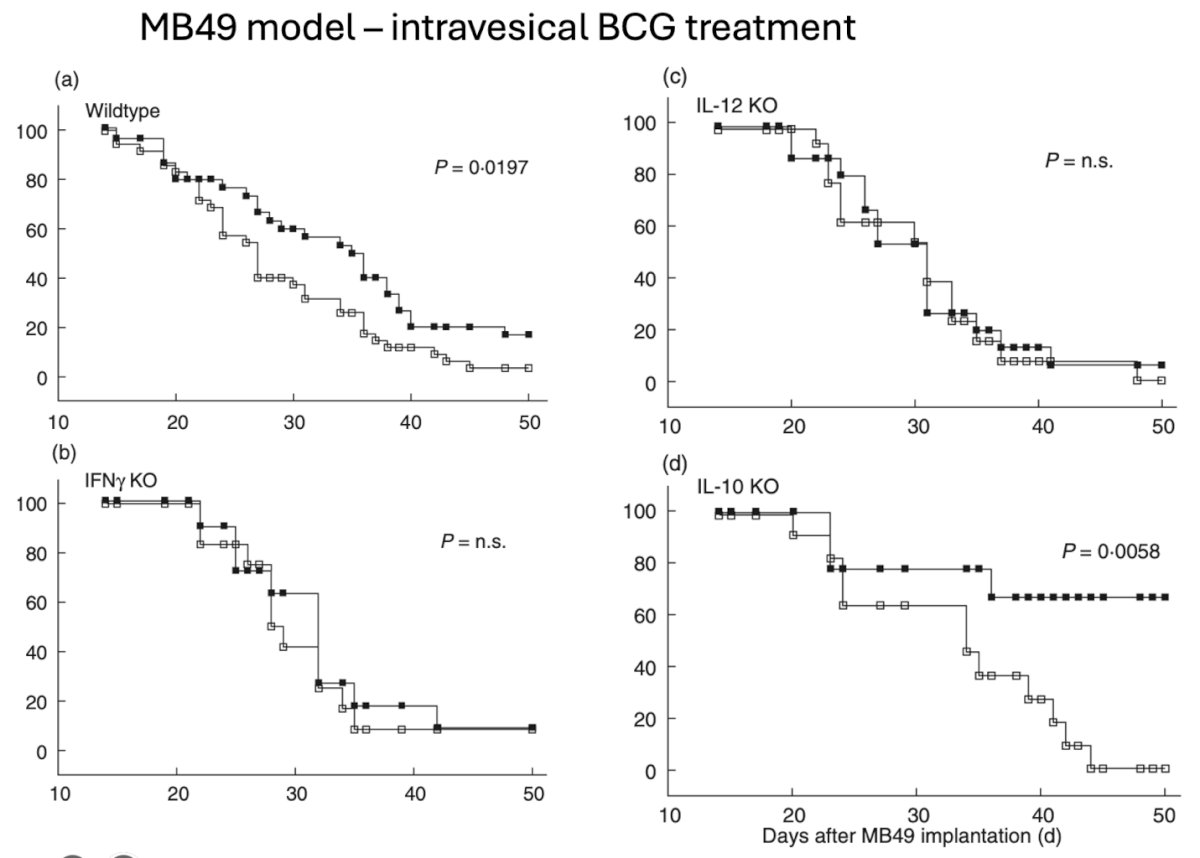
Dr. Garris described data from a preclinical MB49 orthotopic bladder cancer model evaluating the efficacy of intravesical BCG treatment across various immune contexts. As shown below, BCG therapy significantly improved survival in wild-type mice (P = 0.0197). However, in IFNγ knockout and IL-12 knockout mice, both essential components of a type 1 immune response, no survival benefit was observed, underscoring their critical role in mediating BCG's antitumor activity. Interestingly, in IL-10 knockout mice, BCG efficacy was enhanced (P = 0.0058), suggesting that IL-10–a known immunosuppressive cytokine–acts as a negative regulator of response. These findings collectively highlight the importance of a type 1 immune axis and the suppressive role of IL-10 in modulating therapeutic responses to BCG.2
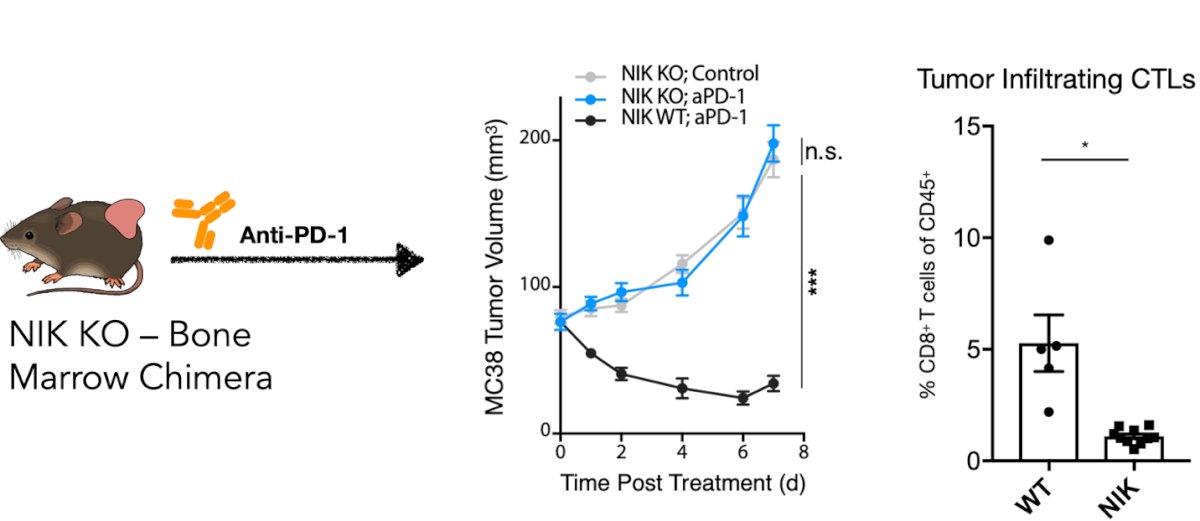
Immunogenic DC maturation is tightly regulated through the non-canonical NF-κB pathway, primarily mediated by CD40 signaling and the NIK-IKKα-RelB axis. CD40 engagement triggers a series of intracellular signals via TRAF2/3, cIAP1, and NIK, culminating in the nuclear translocation of p52/RelB, a key transcriptional complex. Transcriptomic profiling highlights that this pathway is most active in conventional dendritic cell subsets, particularly DC1, as shown by expression of CD40, Map3k14 (NIK), and Relb. Functionally, stimulation with aCD40 or a SMAC mimetic (AZD5582) induces intratumoral IL-12 expression, a hallmark of activated DCs (middle panel in the figure below). However, in NIK knockout mice (bone marrow chimeras), the absence of this signaling impairs anti-PD-1 efficacy and reduces intratumoral CD8⁺ T cell infiltration, underscoring NIK's essential role in shaping the tumor immune microenvironment. Together, these data emphasize the centrality of the CD40-NIK pathway in orchestrating DC immunogenicity and subsequent T cell–mediated tumor rejection.3
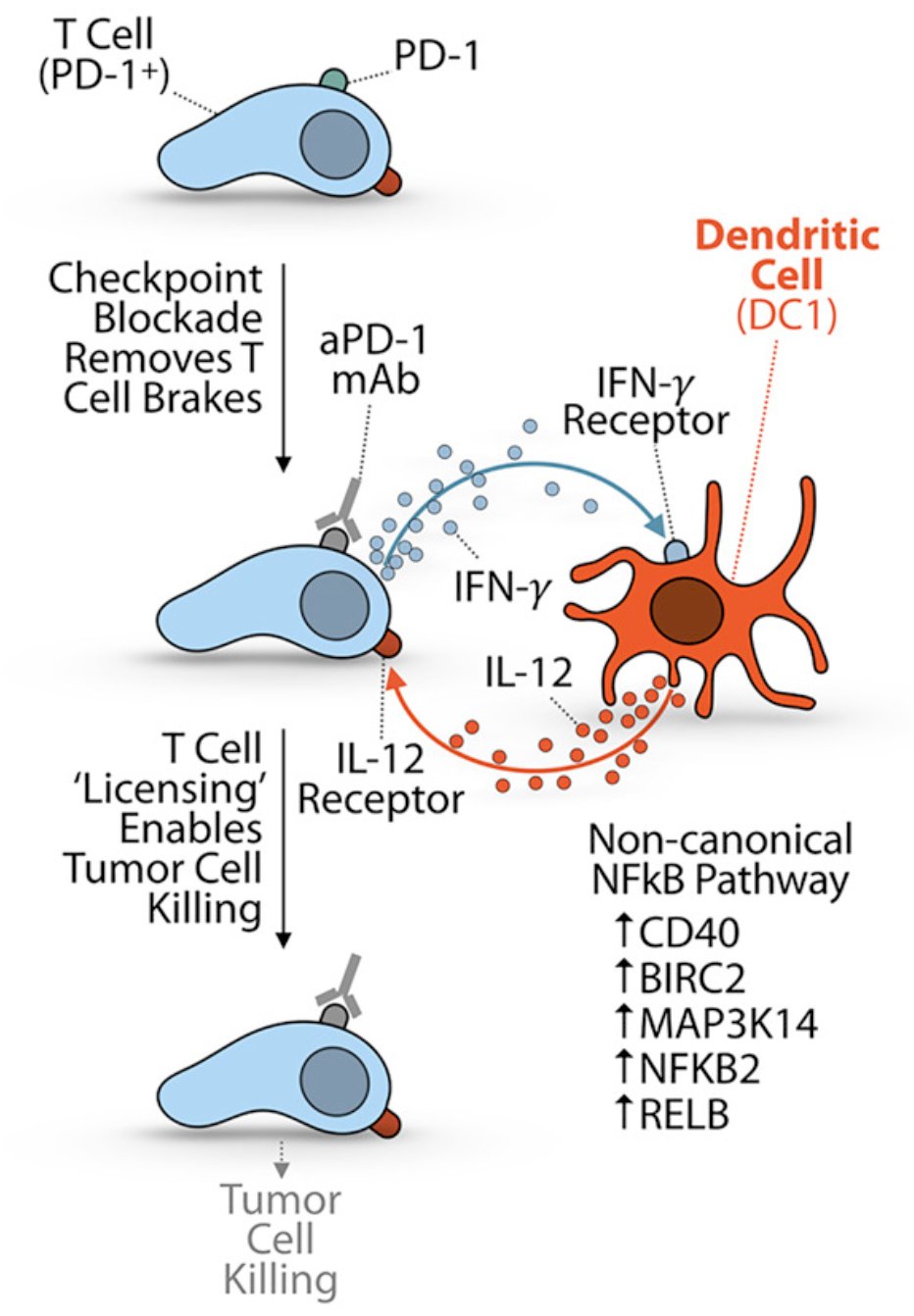
To enhance IL-12 production within the tumor microenvironment, therapeutic strategies aim to engage dendritic cells through two reinforcing mechanisms: checkpoint blockade and non-canonical NF-κB signaling. As illustrated below anti–PD-1 antibodies relieve inhibitory signals on exhausted T cells, enabling them to secrete IFN-γ. This IFN-γ then activates dendritic cells, which, through upregulation of CD40, BIRC2, MAP3K14, NFKB2, and RELB, amplify IL-12 production via the non-canonical NF-κB pathway. IL-12, in turn, licenses CD8⁺ T cells for effective tumor killing, creating a positive feedback loop that boosts cytotoxic immunity and reinforces local immune activation. So, it becomes key to enhance IL-12.
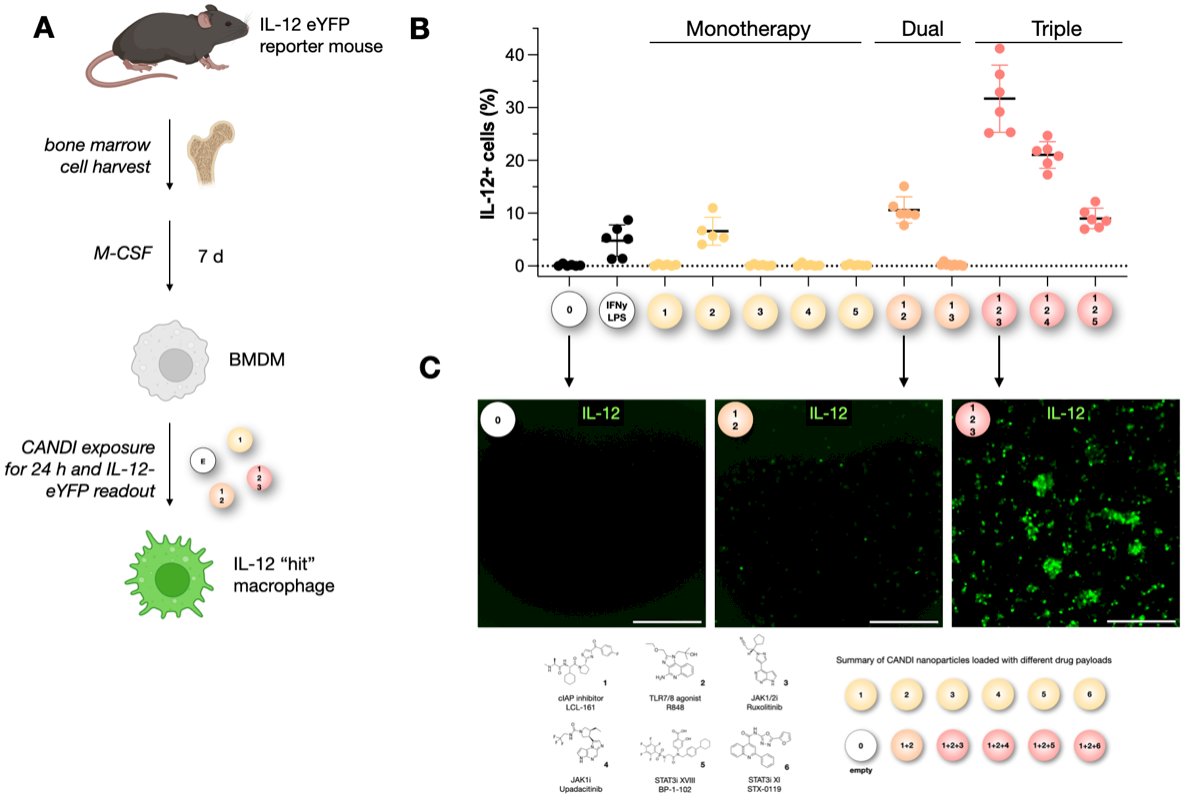
Frederich and colleagues developed a screening platform to identify drug combinations that enhance IL-12 production using IL-12-eYFP reporter mice. Bone marrow–derived macrophages were cultured and exposed to CANDI nanoparticles containing various immune-modulating agents. As shown in panel B, IL-12 induction was minimal with monotherapy, modest with dual therapy, and significantly increased with triple combinations, particularly with agents targeting TLR signaling, JAK/STAT, and non-canonical NF-κB pathways. In the graphic below, there is clear evidence on fluorescence microscopy of a marked increase in IL-12⁺ cells with combination therapies.3
Cyclodextrin-Adjuvant Nanoconstructs Double Inhibitor (CANDI) nanoparticles represent a promising strategy to safely deliver immune stimulants directly to myeloid cells within the tumor microenvironment. Composed of succinyl-β-cyclodextrin and L-lysine, these ~17 nm particles encapsulate immunomodulatory drugs such as cIAP inhibitors, TLR7/8 agonists, and JAK/STAT pathway inhibitors. As shown in the heatmap, CANDI preferentially targets antigen-presenting cells like CD11c⁺ MHCII⁺ dendritic cells and CD11b⁺ macrophages, while sparing lymphocytes and tumor cells. Importantly, hepatotoxicity assays demonstrated that CANDI delivery dramatically reduces systemic toxicity, supporting its translational potential as a safer vehicle for stimulating intratumoral IL-12 production and enhancing anti-tumor immunity.4

Additionally, there is a growing focus on strategies that directly induce IL-12 production within dendritic cells, given their central role in shaping T cell–mediated anti-tumor immunity. In this model, using IL-12 eYFP reporter mice, bone marrow–derived myeloid cells were differentiated into dendritic cells and then exposed to CANDI nanoparticles. While untreated or empty nanoparticle-exposed dendritic cells produced minimal IL-12, stimulation with IFNγ plus LPS, or with CANDI formulations, robustly induced IL-12 production. Notably, the addition of JAK inhibitors significantly enhanced IL-12 expression as illustrated below, suggesting that precise modulation of JAK/STAT signaling pathways can optimize IL-12 induction within dendritic cells, offering a potential mechanism to reinvigorate tumor immune responses.

Dr. Garris highlighted that JAKi boosts DC migration, enhances T cell stimulation, and improves innate immunotherapy against bladder tumors. In-vivo, using an orthotopic MB49 model, mice treated with IV CANDI showed significant reductions in tumor burden, with the CANDI402 group achieving bladder weights nearly identical to healthy controls. This suggests that JAK inhibition amplifies dendritic cell–driven immune activation, resulting in more effective tumor clearance.
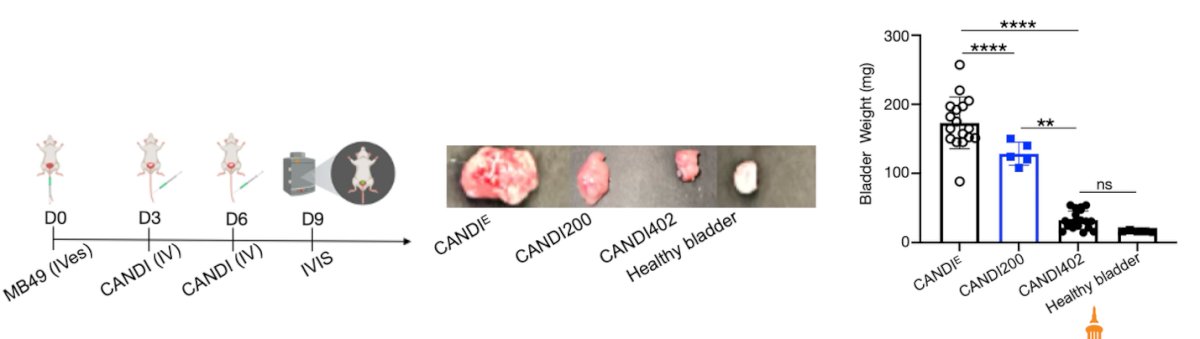
Dr. Garris presented additional evidence that JAK inhibition enhances IL-12–driven immune activation in the tumor-draining lymph nodes (tdLNs) and in the tumor micro-environment (TME). Treatment with CANDI402 significantly increased IL-12⁺ dendritic cells and macrophages compared to both untreated and CANDI200-treated groups, suggesting a more complete activation of adaptive immunity. These findings support that JAKi augments IL-12 production in key APC subsets, promoting a more effective T cell–priming environment.
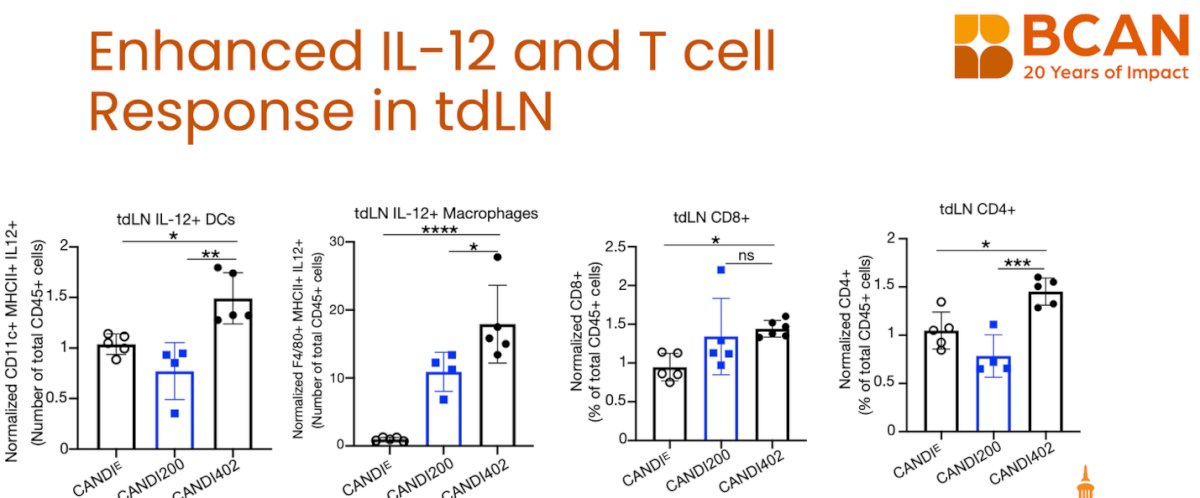
Tumor IL-12 responses require lymphocyte trafficking, as highlighted by Dr. Garris. When lymphocyte movement was blocked with Fingolimod (FTY720), which is used as a treatment for multiple sclerosis. The antitumor effects of CANDI402 were lost, and IL-12–producing cells were not effectively induced in the tumor microenvironment. This underscores that the therapeutic activity of innate immunotherapy relies not only on local delivery but also on active immune cell trafficking from lymphoid tissues to the tumor.

Dr. Garris further explained that lymph node IL-12 responses are impaired by FTY720. Mice treated with FTY720 and CANDI402 showed reduced IL-12⁺ dendritic cells and macrophages in tumor-draining lymph nodes compared to CANDI402 alone. CD4⁺ T cell frequencies were also diminished, while CD8⁺ T cells were not significantly affected. These findings reinforce that lymphocyte trafficking is essential not only for tumor infiltration but also for initiating IL-12–mediated priming within the lymph node compartment.
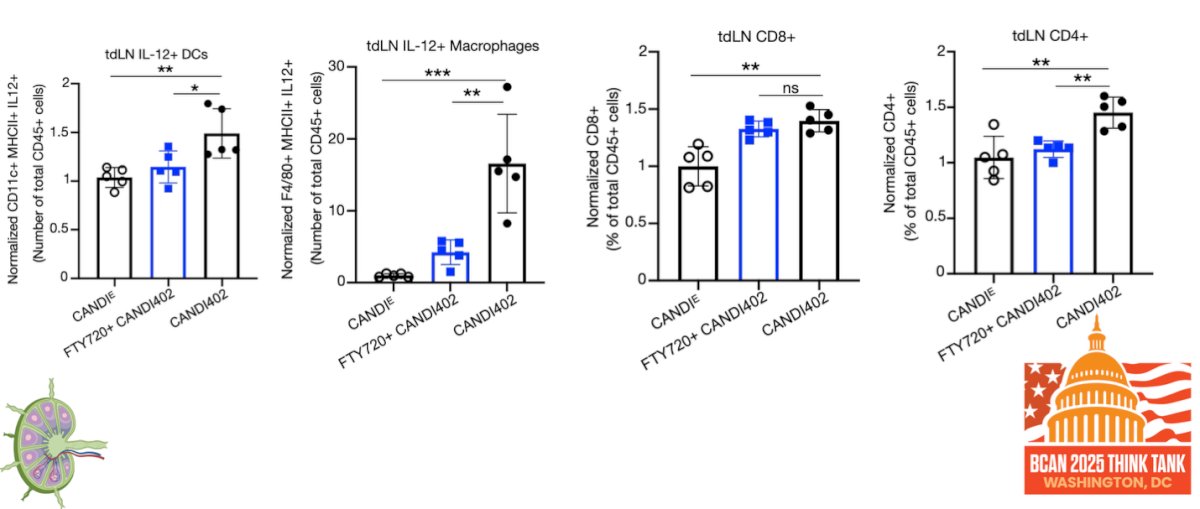
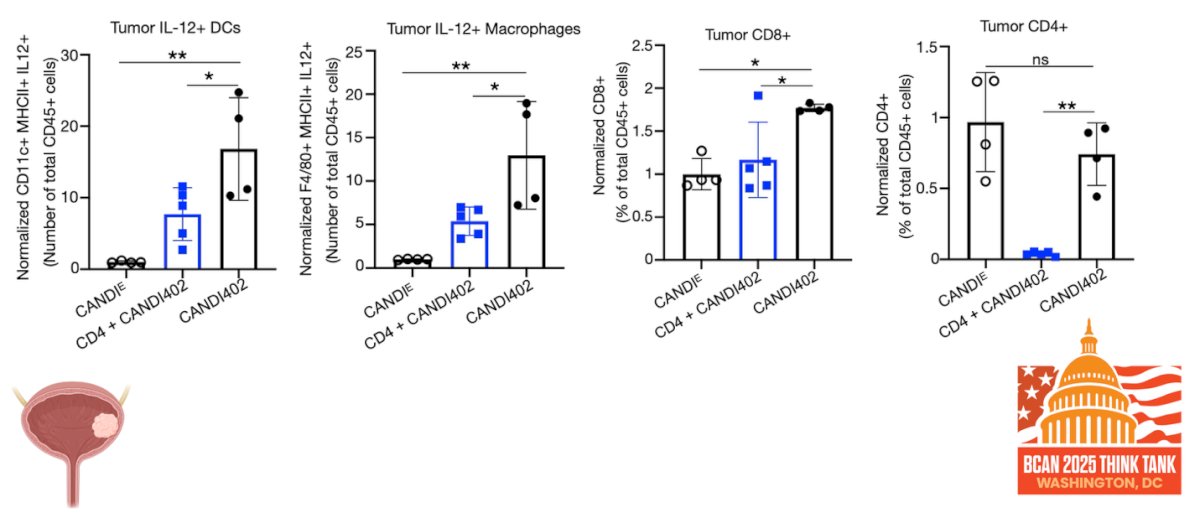
Moreover, CD4 depletion abrogates the effects of innate immunotherapy, highlighting a critical role for CD4⁺ T cells in mediating antitumor activity. As illustrated in the figure below, CD4⁺ T cell presence is necessary to trigger intratumoral IL-12 responses, suggesting that CD4-driven crosstalk is essential for activating myeloid cells and sustaining effective innate immune engagement within the tumor microenvironment.
Dr. Garris highlighted that CANDI innate immunotherapy effectively treats lung metastases, as shown in the figures below. In a metastatic MB49 model, repeated IV dosing with CANDI402 led to a dramatic reduction in tumor burden by Day 18. IVIS imaging confirmed minimal signal in the CANDI402 group compared to widespread lung involvement in untreated controls. Correspondingly, tumor weights were significantly lower in CANDI402-treated mice, and gross pathology showed visibly smaller lung tumors, reinforcing the systemic activity and metastatic efficacy of CANDI-based innate therapy.
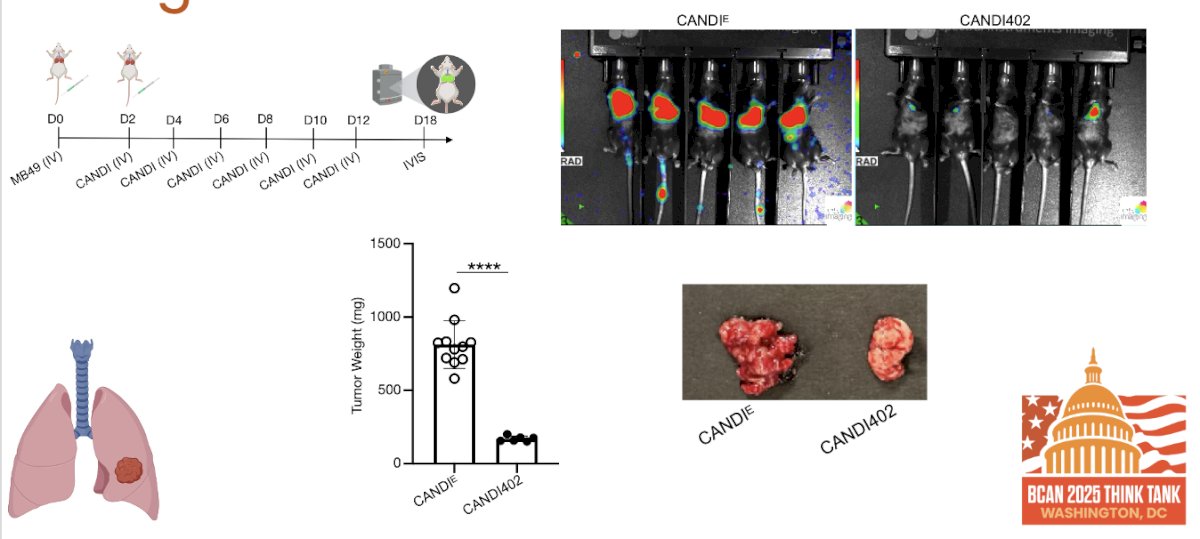
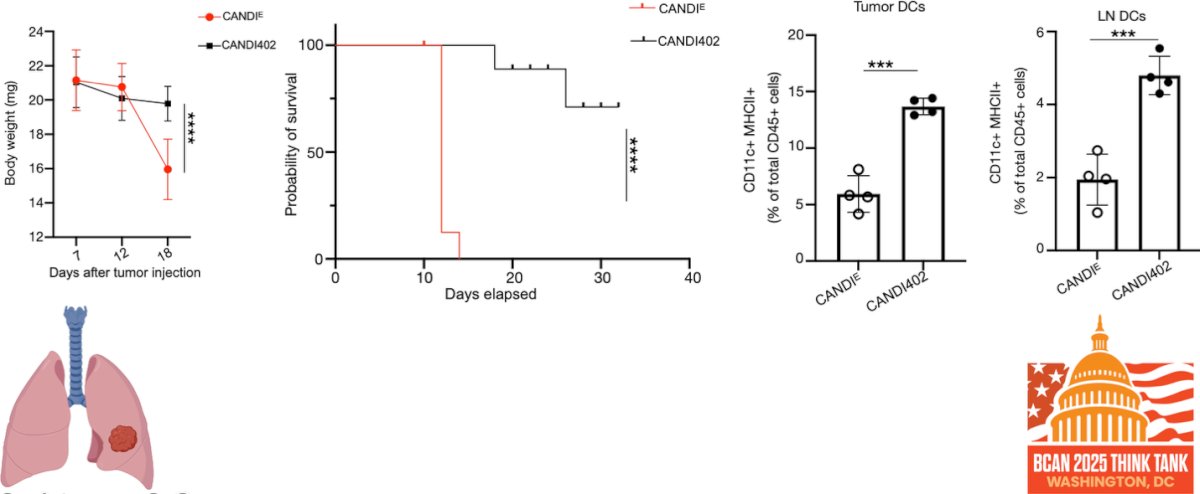
Dr. Garris showed that CANDI402 not only reduces tumor burden but also drives durable antitumor responses. In a lung metastasis model, CANDI402-treated mice demonstrated prolonged survival and had a marked increase in both tumor-infiltrating and lymph node dendritic cells compared to controls. These findings support sustained immune engagement and suggest that innate immunotherapy with CANDI402 may generate long-lasting protection against metastatic disease.
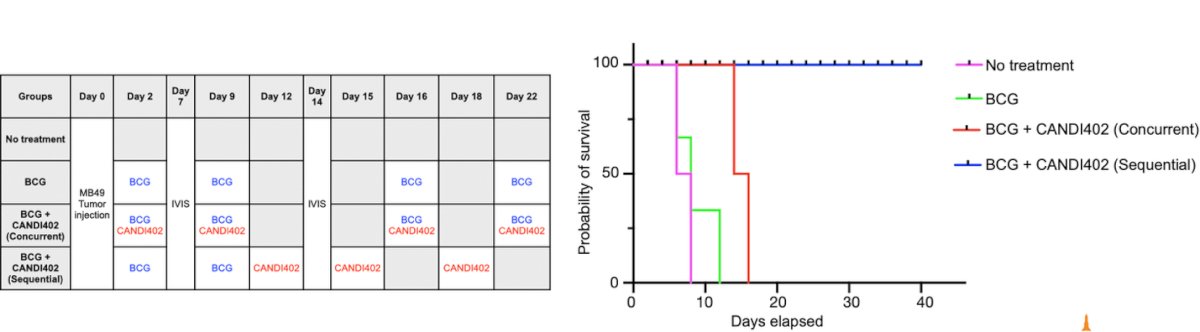
Another potential strategy is the combination of BCG with CANDI immunotherapy, as illustrated below. In this MB49 bladder tumor model, mice receiving sequential BCG followed by CANDI402 showed complete survival and long-term responses, whereas BCG alone, or concurrent administration of both therapies, led to only modest benefit.
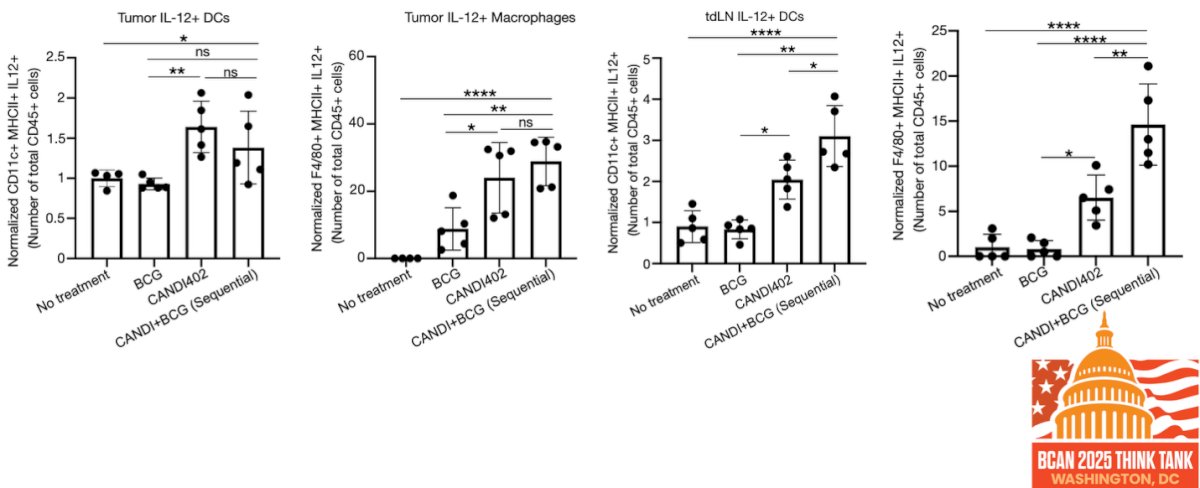
Dr. Garris noted that IL-12 responses induced by treatment varied by strategy. While BCG or CANDI402 alone modestly increased IL-12⁺ dendritic cells and macrophages in both the tumor and draining lymph nodes, the sequential combination of BCG followed by CANDI402 produced again the most robust response across all compartments. This suggests that sequential administration enhances innate immune activation and may be critical for maximizing IL-12–mediated antitumor effects.
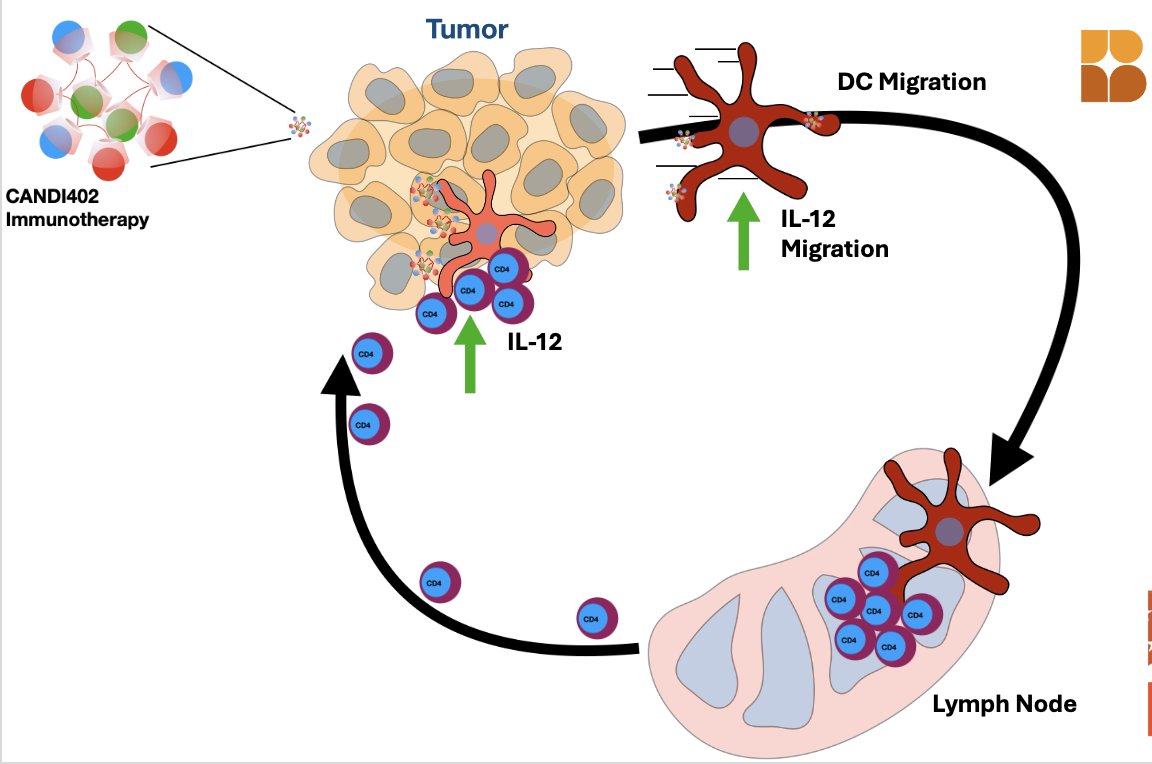
CANDI402 is an innate immunotherapy designed to activate dendritic cells (DCs) and enhance IL-12–driven immune responses against tumors. As illustrated in the figure, CANDI402 stimulates DCs in the lymph node, prompting their migration into the tumor where they interact with CD4⁺ T cells. This interaction triggers IL-12 production, fueling a local immune response. The cycle of DC activation, migration, and CD4⁺ T cell engagement is key to sustaining antitumor activity.
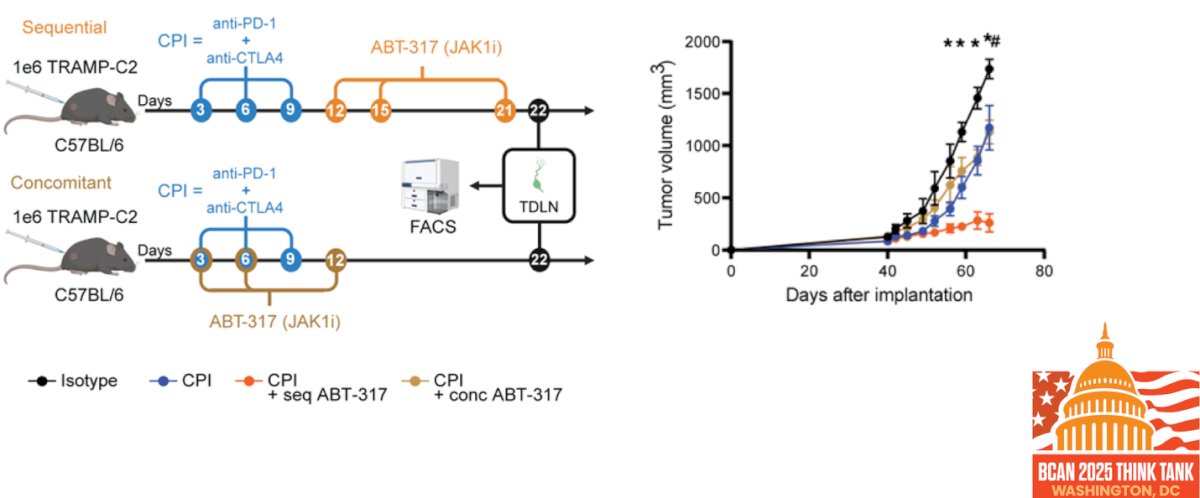
Dr. Garris highlighted findings from Arias-Badia et al. showing that transient blockade of IFN-γ signaling using the JAK1 inhibitor ABT-317 can enhance antitumor T cell responses when paired with checkpoint inhibitors (CPI) in preclinical models. Importantly, only sequential administration of ABT-317 after CPI led to reduced toxicity and significantly better tumor control, as shown by delayed tumor growth. This approach preserved effector T cell function, limited activation-induced T cell death, and promoted memory responses that protected against tumor rechallenge. These results suggest that timed JAK inhibition can overcome immune-intrinsic resistance and improve the durability of CPI responses.5
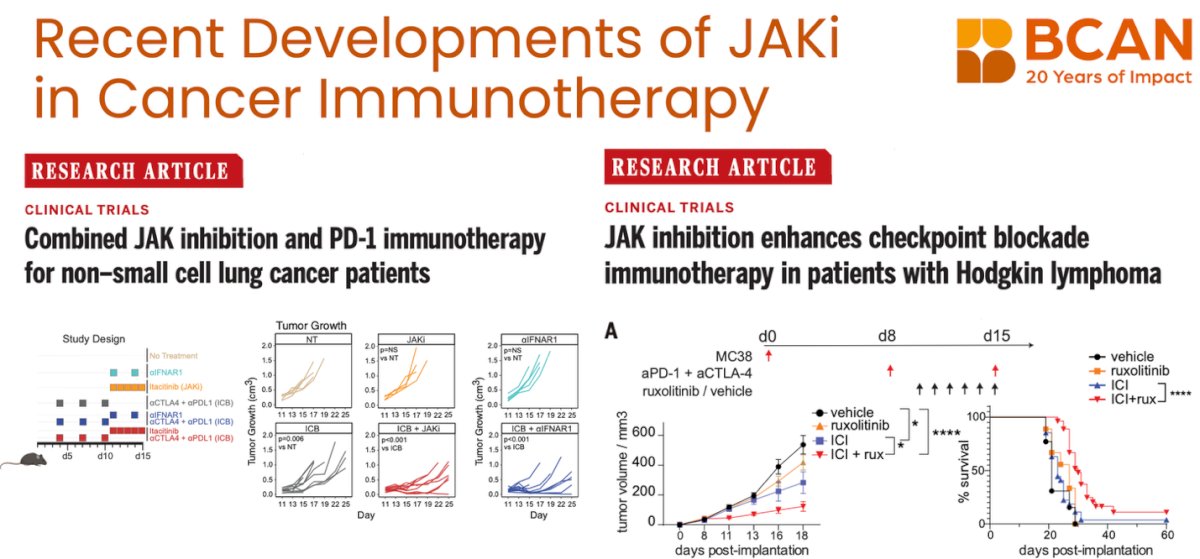
Lastly, Dr. Garris highlighted emerging clinical trials evaluating the combination of JAK inhibition and checkpoint blockade in patients with non-small cell lung cancer and Hodgkin lymphoma. Early results from these studies suggest that targeting JAK signaling may enhance the efficacy of immunotherapy in the clinic, echoing preclinical findings and supporting further exploration of this combinatorial strategy to overcome resistance and improve durable responses
Dr. Garris wrapped up his presentation with the following key takeaways:
- JAKi can boost myeloid stimulation and production of IL-12
- Dendritic cell migration can be augmented with JAKi
- JAKi-boosted innate stimulants targeting myeloid cells can efficiently treat mouse bladder cancer models
- Combination of upfront innate stimulants + JAKi could make agonistic therapy more tolerable
- CD4-driven responses in the TME condition myeloid IL-12
Presented by: Christopher Garris, PhD, Massachusetts General Hospital, Harvard Medical School, Boston, MA
Written by: Julian Chavarriaga, MD – Urologic Oncologist at Cancer Treatment and Research Center (CTIC) via Society of Urologic Oncology (SUO) Fellow at The University of Toronto. @chavarriagaj on Twitter during the Bladder Cancer Advocacy Network (BCAN) Bladder Cancer Think Tank 2025 held in Washington D.C., United States, between July 30th and August 1st.
Reference:
- Chen DS, Mellman I. Oncology meets immunology: the cancer-immunity cycle. Immunity. 2013 Jul 25;39(1):1-10. doi: 10.1016/j.immuni.2013.07.012. PMID: 23890059.
- Riemensberger J, Böhle A, Brandau S. IFN-gamma and IL-12 but not IL-10 are required for local tumour surveillance in a syngeneic model of orthotopic bladder cancer. Clin Exp Immunol. 2002 Jan;127(1):20-6. doi: 10.1046/j.1365-2249.2002.01734.x. PMID: 11882028; PMCID: PMC1906285.
- Garris CS, Arlauckas SP, Kohler RH, Trefny MP, Garren S, Piot C, Engblom C, Pfirschke C, Siwicki M, Gungabeesoon J, Freeman GJ, Warren SE, Ong S, Browning E, Twitty CG, Pierce RH, Le MH, Algazi AP, Daud AI, Pai SI, Zippelius A, Weissleder R, Pittet MJ. Successful Anti-PD-1 Cancer Immunotherapy Requires T Cell-Dendritic Cell Crosstalk Involving the Cytokines IFN-γ and IL-12. Immunity. 2018 Dec 18;49(6):1148-1161.e7. doi: 10.1016/j.immuni.2018.09.024. Epub 2018 Dec 11. Erratum in: Immunity. 2022 Sep 13;55(9):1749. doi: 10.1016/j.immuni.2022.07.021. PMID: 30552023; PMCID: PMC6301092.
- Kim HS, Halabi EA, Enbergs N, Kohler RH, Fei F, Garris CS, Weissleder R. A non-lipid nucleic acid delivery vector with dendritic cell tropism and stimulation. Theranostics. 2024 May 5;14(7):2934-2945. doi: 10.7150/thno.95267. PMID: 38773971; PMCID: PMC11103498.
- Arias-Badia M, Chen P, Lwin YM, Srinath A, Lyu A, Fan Z, Kwek SS, Luong DN, Setayesh A, Sakamoto M, Clark M, Lea A, Wolters RM, Goodearl A, Harding FA, Gorman JV, Ritacco W, Fong L. Sequential JAK inhibition enhances antitumor immunity after combined anti-PD-1 and anti-CTLA4. JCI Insight. 2025 Feb 27;10(7):e187921. doi: 10.1172/jci.insight.187921. PMID: 40014402; PMCID: PMC11981626.